CSB project students present their year’s work in research
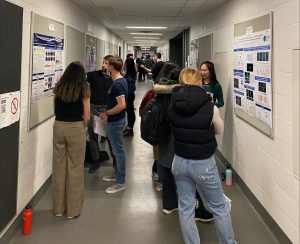 A year of research by CSB project students culminated in a poster session on March 31st, 2023. The students used the posters to…
A year of research by CSB project students culminated in a poster session on March 31st, 2023. The students used the posters to…
 Amanda Miles’ PhD studies in Prof…
Amanda Miles’ PhD studies in Prof…Congratulations to Prof. Ashley Bruce and Prof. Alan Moses on their promotion to Full Professor
It is our pleasure to announce that Ashley Bruce and Alan Moses have been promoted to Full Professor in the Department of Cell and Systems Biology as of July 1, 2019.
